In-Fusion® prawdziwy kombajn w Twoim Laboratorium!
24 Lipca 2024 (Ostatnia aktualizacja 17.10.2024)

Prężny rozwój technik genetyki, biofizyki i biochemii przypadający na lata 70 XX w., pozwolił na powstaje nowej dyscypliny badawczej zwanej biologią molekularną. Jedną z technik, która zrewolucjonizowała pracę z materiałem genetycznym była opracowana w 1973 roku pierwsza metoda łączenia sekwencji DNA, której funkcjonowanie oparte było o trawienie restrykcyjne i ligację (Cohen i wsp., 1973). Stworzony w ten sposób syntetyczny bakteryjny plazmid został z powodzeniem wykorzystany do przeprowadzenia klonowania molekularnego (ang. molecular cloning). W istocie metoda ta polega na odpowiednim przygotowaniu interesującej sekwencji, wprowadzeniu jej do komórki gospodarza z zastosowaniem odpowiedniego nośnika (wektora), a następnie jej powieleniu.
Jak wspomniano, konwencjonalne metody klonowania molekularnego bazują na enzymach restrykcyjnych (tzw. restryktazach), które rozpoznają specyficzne sekwencje DNA długości od kilku do kilkunastu nukleotydów (zwykle 4–8 pz). Rozpoznanie charakterystycznej dla danego enzymu sekwencji tzw. miejsca restrykcyjnego, powoduje hydrolizę wiązań fosfodiestrowych (najczęściej w obrębie sekwencji rozpoznawanej przez enzym) czego konsekwencją jest powstanie tzw. „lepkich” bądź „tępych” końców (odpowiednio, sticky ends oraz blunt ends). Odpowiednio przygotowany, klonowany fragment DNA oraz jego nośnik (wektor) są następnie łączone z wykorzystaniem ligazy i wprowadzane do organizmu biorcy.
Ta metoda klonowania jest bardzo wszechstronna i odegrała kluczową rolę w dziedzinie genetyki, biotechnologii i medycyny, umożliwiając rozwój organizmów modyfikowanych genetycznie, terapii genowych i produkcji farmaceutyków. Niesie ona jednak ze sobą szereg ograniczeń, do najważniejszych z nich niewątpliwie należą:
⦁ zależność od sekwencji rozpoznawanych przez enzymy restrykcyjne;
⦁ ograniczona ilość możliwych do zastosowania enzymów restrykcyjnych wynikająca np.: z występowania miejsca restrykcyjnego w obrębie klonowanej sekwencji;
⦁ duże prawdopodobieństwo zmiany ramki odczytu etc.
Współczesne metody łączenia fragmentów DNA w warunkach in vitro, które bazują na homologii terminalnych odcinków łączonych sekwencji, eliminują lub znacząco zmniejszają ograniczenia charakteryzujące konwencjonalne metody oparte o enzymy restrykcyjne. Ze względu na fakt, iż metody te nie wprowadzają dodatkowych nukleotydów, tym samym nie pozostawiając śladu po łączeniu fragmentów DNA, nazywane są również „bezszwowymi” (ang. Seamless). Dotychczas w literaturze opisanych zostało wiele metod bazujących na homologii sekwencji, jak m.in.: Overlap Extension Polymerase Chain Reaction (OE-PCR), Circular Polymerase Extension Cloning (CPEC)(Quan oraz Tian, 2009), Sequence and Ligation-independent Cloning (SLIC)(Li oraz Elledge, 2007), oraz Seamless Ligation Cloning Extract (SLiCE)(Zhang i wsp., 2012). Jednakże ze względu na prostotę, relatywnie niskie koszty reakcji oraz wysoką wydajność łączenia fragmentów DNA, aktualnie największą popularnością cieszą się jedynie dwie metody.
Pierwszą z nich jest metoda opracowana w 2009 roku przez Gibson i wsp. (2009), która została nazwana od nazwiska jej odkrywcy. Główną zaletą metody jest fakt, iż łączone fragmenty DNA są poddawane jednoczesnemu działaniu trzech enzymów w jednej mieszaninie reakcyjnej. W początkowym etapie łączone fragmenty DNA amplifikuje się z jednoczesnym wprowadzeniem nakładających się, flankujących sekwencji homologicznych (długość 15-20 nt). Tak otrzymane, oczyszczone produkty miesza się w odpowiednich stężeniach i poddaje działaniu mieszaniny zawierającej egzonukleazę, polimerazę DNA oraz termostabilną ligazę. Egzonukleaza o aktywności 5’→3’ generuje jednoniciowe homologiczne lepkie końce, polimeraza DNA (o ograniczonej aktywności proofreading) uzupełnia luki pomiędzy elementami konstruktu, a termostabilna ligaza scala wszystko w całość.
Nieco odświerzoną wariacją metody Gibson jest metoda NEBuilder HiFi DNA Assembly, która wykorzystuje tą samą technologię, ale znajdująca się w premiksie polimeraza jest enzymem o wysokiej wierności. Znacząco poprawia to jakość powstających konstruktów (mniej błędnie wprowadzonych nukleotydów w obszarze homologicznych fragmentów). W obu przypadkach termostabilna ligaza jest obecna w mieszaninie reakcyjnej od samego początku trwania reakcji. Konsekwencją tego jest realatywnie wysoka wydajność „zamykania się” pustych plazmidów, co z kolei znacząco mniejsza efektywność klonowania molekularnego.
Opracowana w 2012 roku technologia In-Fusion® (Irwin i wsp., 2012) zrewolucjonizowała działania związane z łączeniem fragmentów DNA o zróżnicowanej długości. Podobnie jak metoda Gibson, reakcja In-Fusion® również przebiega w jednej probówce i jest jednoetapowa. W tym przypadku zostasowano jednak zauważalnie szybszą egzonykleazę o aktywności 3’→5’ co pozwoliło na skrócenie czasu reakcji do 15 minut. Długość łączonych, homologicznych fragmentów DNA zawsze wynosi 15 nukleotydów co niweluuje konieczność optymalizacji warunków rakcji. Jednakże najważniejszym udoskonaleniem technologii In-Fusion® jest fakt, iż mieszanina reakcyjna pozbawiona jest enzymu o aktywności ligazy. Łączenie fragmentów DNA i składanie kompletnych plazmidów realizowane jest z wykorzystaniem naturalnej „maszynerii” enzymatycznej gospodarza (in vivo). Niemal całkowicie wyeliminowało to problem tła reakcji, będącego konsekwencją zamykania się pustych plazmidów. Ograniczyło to również wprowadzanie błędnych nukleotydów i tworzenie niepoprawnych konstruktów.
Uproszczony schemat reakcji In-Fusion® przedstawiono na Figurze 1.
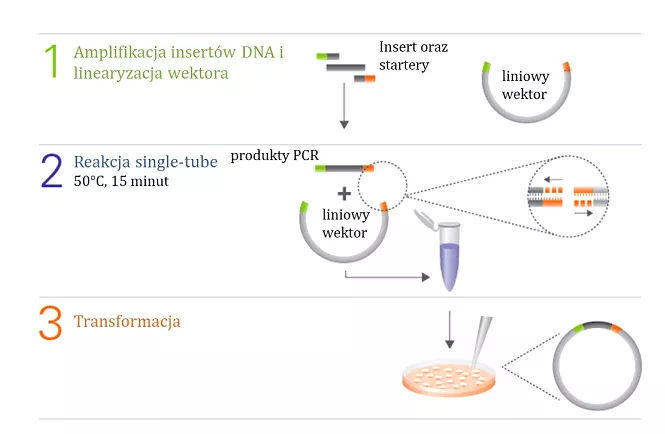
Fig. 1. Schemat reakcji In-Fusion®[https://www.takarabio.com/learning-centers/cloning/in-fusion-cloning-general-information/in-fusion-cloning-overview].
Ponadto, oferowane przez TakaraBio zestawy In-Fusion® to prawdziwe MULTIZADANIOWE KOMBAJNY do zadań specjalnych. Reakcja In-Fusion® może być z powodzeniem zastosowana m.in.:
⦁ do klonowania pojedynczych jak i wielofragmentowych konstruktów z wydajnością wynoszącą nawet 95%;
⦁ insercji, delecji lub substytucji określonych sekwencji kodu genetycznego, co znajduje zastosowanie m.in. w ukierunkowanej mutagenezie;
⦁ do klonowania regionów zmiennych przeciwciał.
Są to jedynie nieliczne z zastosowań, które w laboratoriach na całym świecie znajdują zestawy In-Fusion®. Niektóre z nich zestawiono na Figurze 2.
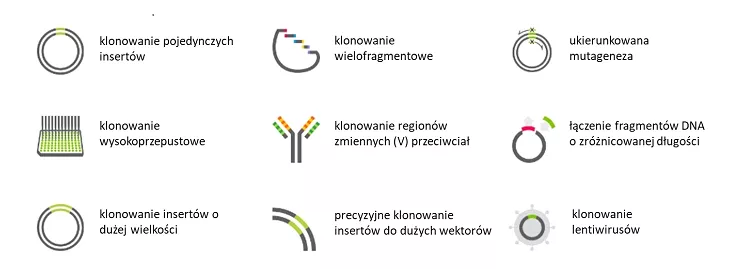
Fig. 2. Przykładowe zastosowania technologii In-Fusion®[https://www.takarabio.com/learning-centers/cloning/in-fusion-cloning-general-information/in-fusion-cloning-overview].
Specjaliści TakaraBio przeprowadzili szereg doświadczeń mających na celu porównanie wydajności i efektywności metody Gibson oraz technologii In-Fusion®. Wyniki przedstawione na Figurze 3A jednoznacznie wskazują, że zastosowanie In-Fusion® do wielofragmentowego klonowania, aż 5 insertów o wielkości od 405 do 1005 pz, pozwoliło na uzyskanie zdecydowanie większej ilości kolonii transformowanych bakterii, przy jednocześnie bardzo wysokiej 90% dokładności klonowania, w porównaniu z reakcją prowadzoną z wykorzystaniem metody Gibson. Podobną zależność zaobserwowano dla klonowania pojedynczego, acz bardzo dużego (34,2 kpz) fragmentu DNA (Figura 3B).
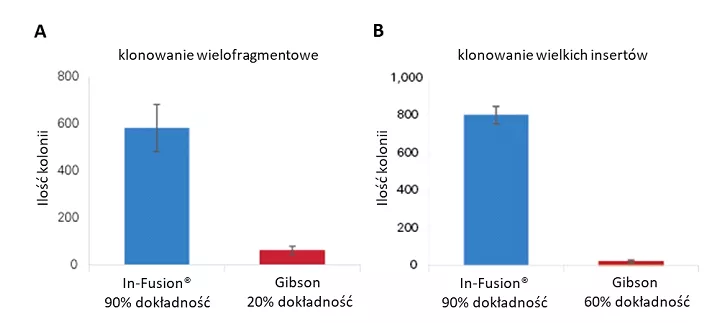
Fig. 3. Porównanie wydajności pomiędzy In-Fusion® Snap Assembly i Gibson Assembly: A) gdy pięć fragmentów (405–1005 pz) wklonowano jednocześnie do wektora pUC19 (2,7 kpz).
Dokładność określono metodą sekwencjonowania Sangera; B) gdy duży fragment o wielkości 34,2 kpz wklonowano do wektora pMET (2,6 kb). Dokładność określono metodą PCR na koloniach; Przedstawione na wykresach wartości są średnimi z trzech niezależnych reakcji klonowania. Słupki błędów pokazują ± odchylenia standardowe [https://www.takarabio.com/learning-centers/cloning/in-fusion-cloning-general-information/in-fusion-cloning-and-competition/in-fusion-snap-assembly-vs-geneart-gibson-assembly-hifi].
Podobne badania przeprowadzono również celem porównania technologii In-Fusion® Snap Assembly i metody NEBuilder HiFi DNA Assembly. W tym przypadku do zlinearyzowanego metodą Inverse PCR wektora wklonowano pojedyńcze inserty o wielklości 3,8 kpz oraz 34,2 kpz. W obu przypadkach zastosowanie technologii In-Fusion® pozwoliło na uzyskanie dwukrotnie większej ilości transformantów, w porównaniu z wynikami uzyskanymi dla NEBuilder HiFi DNA Assembly. Dla reakcji prowadzonej z wykorzystaniem In-Fusion®, aż 95% spośród uzyskanych transformantów charakteryzowało się poprawną sekwencją.
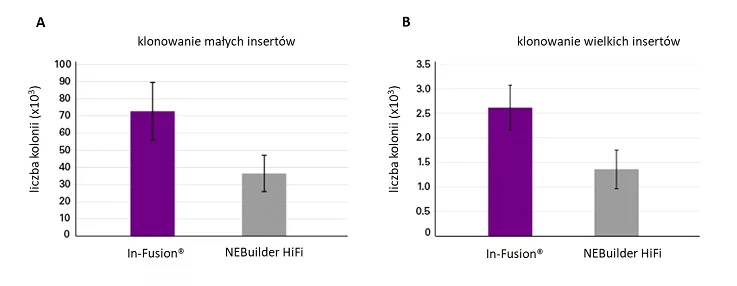
Fig. 4. Porównanie wydajności technologii In-Fusion® i NEBuilder HiFi przy użyciu inverse PCR.
Pojedynczą wstawkę o wielkości 3,8 kpz (panel A) lub wstawkę adenowirusa o wielkości 34,2 kpz (panel B) wklonowano do wektora o wielkości 2,7 kpz, który linearyzowano metodą inverse PCR. Reakcje klonowania przeprowadzono w trzech powtórzeniach. Przebieg reakcji zaprojektowano zgodnie ze specyfikacjami producentów. Po transformacji i wysianiu, 20 kolonii z każdego powtórzenia analizowano metodą sekwencjonowania Sangera (dla wstawki 3,8 kpz) lub colony PCR (dla wstawki adenowirusa) w celu określenia dokładności klonowania [https://www.takarabio.com/learning-centers/cloning/in-fusion-cloning-general-information/in-fusion-cloning-and-competition/in-fusion-snap-assembly-vs-nebuilder-hifi].
Analogiczne badania przeprowadzono dla wektorów linearyzowanych z wykorzystaniem enzymów restrykcyjnych. W tym przypadku In-Fusion® Snap Assembly również okazał się bardziej skuteczny i pozwolił na otrzymanie 5x większej ilości kolonii dla fragmentów z 5’-lepkimi końcami, 8x większej ilości kolonii dla fragmentów z tępymi końcami oraz aż 16x większej ilości kolonii dla fragmentów z 3’-lepkimi końcami, w porównaniu z wynikami uzyskanymi dla NEBuilder HiFi DNA Assembly (wyniki nie pokazane, dostępne na https://www.takarabio.com/learning-centers/cloning/in-fusion-cloning-general-information/in-fusion-cloning-and-competition/in-fusion-snap-assembly-vs-nebuilder-hifi).
Poniżej wyszczególnione są warianty, w których zakupić można zestaw do klonowania In-Fusion®. Przygotowane przez TakaraBio zestawy wychodzą naprzeciw oczekiwaniom klientów i pozwalają na zakup produktów wzbogaconych m.in.: o komórki kompetentne, kolumienki do oczyszczania produktów PCR oraz w postaci gotowych premiksów w formie zliofilizowanej.
- In-Fusion® Snap Assembly Master Mix – zestawy od 10 do 1000 reakcji (638947; 638948; 638949; 638949; 638943; 638944);
- In-Fusion® Snap Assembly Starter Bundle – NucleoSpin Gel and PCR Clean-Up; Stellar Competent Cells; PrimeSTAR Max DNA Polymerase – 10 reakcji (638945);
- In-Fusion® Snap Assembly Value Bundle – NucleoSpin Gel and PCR Clean-Up; Stellar Competent Cells; PrimeSTAR Max DNA Polymerase – 50 reakcji (638946);
- In-Fusion® Snap Assembly Master Mix with Competent Cells – Stellar Competent Cells – zestawy od 10 do 50 reakcji (638951; 638952);
- In-Fusion® Snap Assembly EcoDry™ Master Mix – zestawy w formie zliofilizowanej w stripach PCR – zestawy od 8 do 96 reakcji (638954; 638955; 638956);
- In-Fusion® Snap Assembly EcoDry™ Master Mix with Competent Cells – zestawy w formie zliofilizowanej w stripach PCR w zestawie z komórkami Stellar Competent Cells – 96 reakcji (638953).
W Laboratorium jak na żniwach, najlepiej sprawdzają się MULTIZADANIOWE KOMBAJNY do zadań specjalnych!
WYPRÓBUJ TECHNOLOGIE In-Fusion® W SWOIM LABORATORIUM
Cohen, S. N., Chang, A. C. Y., Boyer, H. W., and Helling, R. B. (1973). Construction of biologically functional bacterial plasmids in vitro. Proc. Natl. Acad. Sci. U. S. A. 70, 3240–3244. doi:10.1073/pnas.70.11.3240.
Quan, J., and Tian, J. (2009). Circular polymerase extension cloning of complex gene libraries and pathways. PLoS One 4, 1–6. doi:10.1371/journal.pone.0006441.
Li, M. Z., and Elledge, S. J. (2007). Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods 4, 251–256. doi:10.1038/nmeth1010.
Zhang, Y., Werling, U., and Edelmann, W. (2012). SLiCE: A novel bacterial cell extract-based DNA cloning method. Nucleic Acids Res. 40, 1–10. doi:10.1093/nar/gkr1288.
Gibson, D. G., Young, L., Chuang, R. Y., Venter, J. C., Hutchison, C. A., and Smith, H. O. (2009). Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 6, 343–345. doi:10.1038/nmeth.1318.
Chad R. Irwin, Ch. R., Farmer, A., Willer, D. O. and Evans, D. H. (2012). In-Fusion® Cloning with Vaccinia Virus DNA Polymerase. Methods in Molecular Biology, volume 890, 23-35.





