Primescript
08 Marca 2023 (Ostatnia aktualizacja 22.10.2024)
Koniec z ustępstwami!
Odwrotna transkrypcja z PrimeScript™ RT
To unikalny enzym MMLV działający w 37°C i posiadający zdolność do rozplatania wiązań wodorowych w obrębie cząsteczki RNA tzw. strand displacement.
Cząsteczka RNA to jednoniciowa struktura zbudowana z nukleotydów, z odsłoniętymi zasadami azotowymi. Oddziałują one ze sobą komplementarnie, przez co cząsteczki RNA mają nieraz bardzo skomplikowaną strukturę i pełnią rolę szkieletową np. w rybozymach i bardziej skomplikowanych strukturach jak rybosomy.
Zwykle, aby przepisać RNA na stabilniejsze cDNA, wykorzystywana jest wysoka temp. w trakcie przepisywania lub procedura wymaga dodatkowego kroku wstępnej denaturacji próbki.
W wyniku podgrzewania pękają wiązania wodorowe w obrębie cząsteczki. Enzym ma dostęp do matrycy oraz mogą się przyłączyć primery wyznaczające początek syntezy. To szczególnie ważne, gdy wykorzystywane są primery genospecyficzne dłuższe od standardowych losowych heksametrów czy oligo dT.
Niestety wysoka temp. działa nie tylko na wiązania wodorowe, ale także niszcząco na całe cząsteczki RNA, powodując ich degradację – liza termiczna.
Dobrze przedstawiają ją doświadczenia poniżej, przepisania drabinki RNA na primery oligo dT oraz rRNA na primery genospecyficzne (Fig2). W pierwszym przypadku (Fig1) odwrotna transkrypcja zaczynała się od ogona poliA i trwała do końca matrycy lub napotkania przez enzym struktury wyższego rzędu. Gdy do tego dochodziło zamiast pojedynczego prążka możemy zaobserwować tzw. „smear”. W przypadku gdy RNA było przepisywane przez PrimeScript™ w niskiej temp. nie obserwujemy tak znaczącego zaciemnienia.
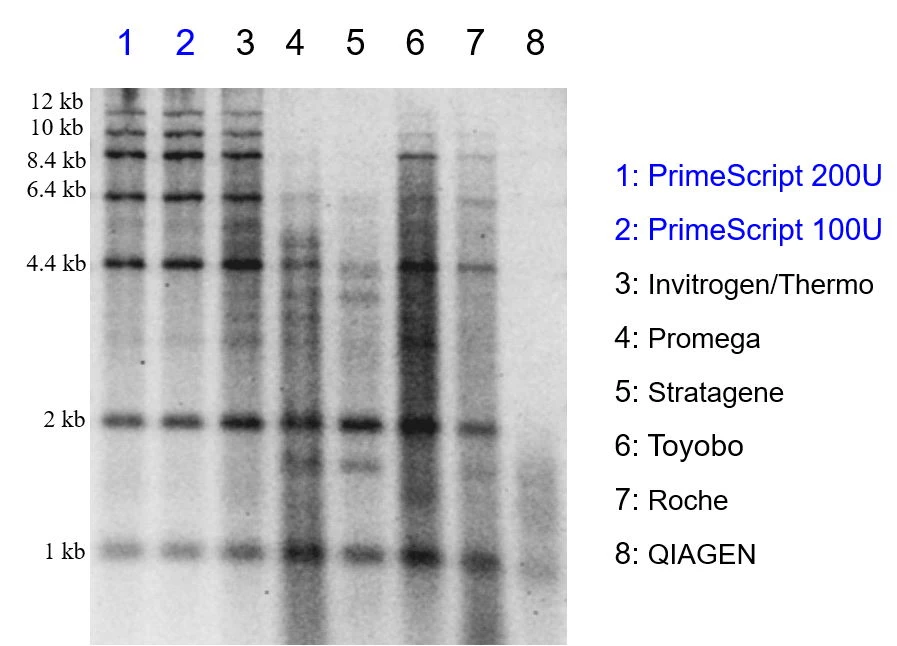
Fig.1 PrimeScript™ - specyficzność i jakość transkrypcji
Zdjęcie przedstawia wyniki rozdziału przepisanej na primery oligo dT drabinki RNA (1; 2; 4,4; 6,4; 8,4; 10; 12 kb). Reakcje były przeprowadzone zgodnie z zalecanymi przez producentów protokołami. Na żel alkaliczny nakładano próbki o takim samym stężeniu. W celu uzyskania większej czułości żel wybarwiono SYBR Green. Produkty uzyskane po transkrypcji za pomocą PrimeScript™ (studzienki 1 i 2) były lepszej jakości i rozdzielczości niż te uzyskane przez porównywane enzymy. Powstałe produkty były specyficzne, pozyskane z dobrą wydajnością i przy wygenerowaniu niewielkiej ilości tła.
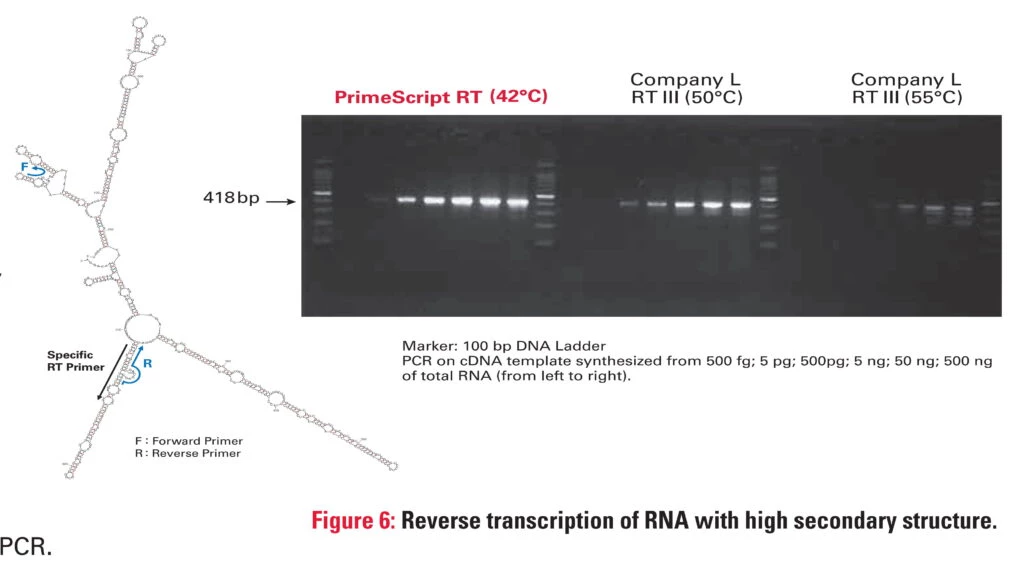
* Temp. reakcji transkrypcji z genospecyficznym primerem
Fig.2 PrimeScript™ zapewnia INTEGRALNOŚĆ RNA. Przedstawione są wyniki procesu rt-PCR cząsteczki rRNA dla podjednostki rybosomu 28S. W pierwszym etapie z puli całkowitego RNA w różnych stężeniach przepisano rRNA w oparciu o genospecyficzny primer (a.). W tym celu użyto enzymów pracujących w różnych temp. Następnie przeprowadzono PCR generując 418 nt fragment rozdzielany na żelu (end-point). Doświadczenie dokumentuje, że wraz ze wzrostem temp. zmniejsza się stabilność RNA. Im wyższa temp. odwrotnej transkrypcji, tym bardziej zmniejsza się integralność RNA. Czyli spada ilość matrycy z której wygenerowane cDNA posłuży w kolejnym etapie do amplifikacji. Zmniejszenie integralności RNA skutkuje słabszym, niejednorodnym prążkiem. Użycie primera genospecyficznego dobrze uwidacznia opisane zjawisko. Podwyższanie temp. reakcji lub wstępna denaturacja służą zniszczeniu struktury cząsteczki, aby wydajnie przepisać RNA na cDNA. Są one jednak pozornie niezbędne i mają konsekwencje w wydajności reakcji, jej czułości i specyficzności. PrimeScript™ dzięki zdolnościom do rozplatania nici nie wymaga wysokiej temp. do wydajnego przepisania RNA (b.).
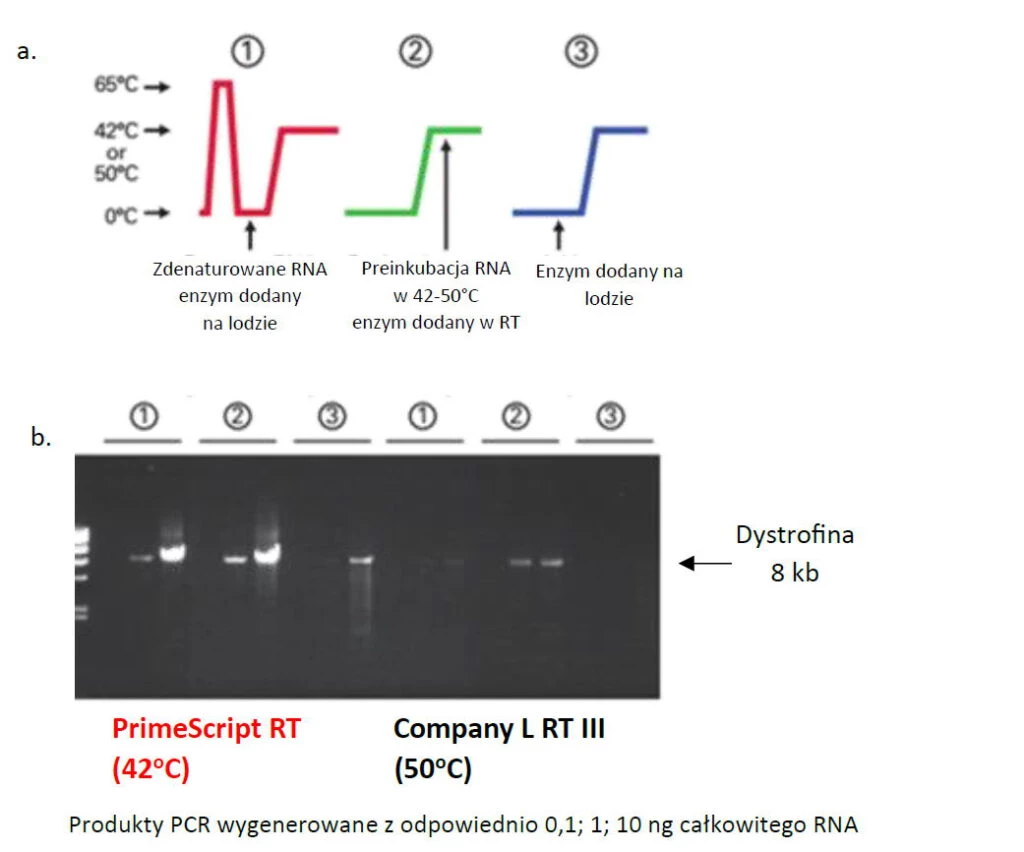
Fig.3. Nie zależnie od momentu dodania, PrimeScript™ znacznie wydajniej przepisał mRNA dystrofiny od porównywanego enzymu. Bez etapu denaturacji (3) konkurencyjna transkryptaza nie podjęła syntezy.
PrimeScript™ dzięki wspomnianym właściwością do rozplatania matrycy nie wymaga podwyższenia temp. aby wniknąć pomiędzy nici w syntezować pełnej długości cDNA. Nie zależnie od momentu dodania, wykazuje dobrą efektywność syntezy (Fig.3). Dzięki temu możliwe jest przepisanie wymagającego RNA złożonego nawet w 75% z par GC (Fig.4).
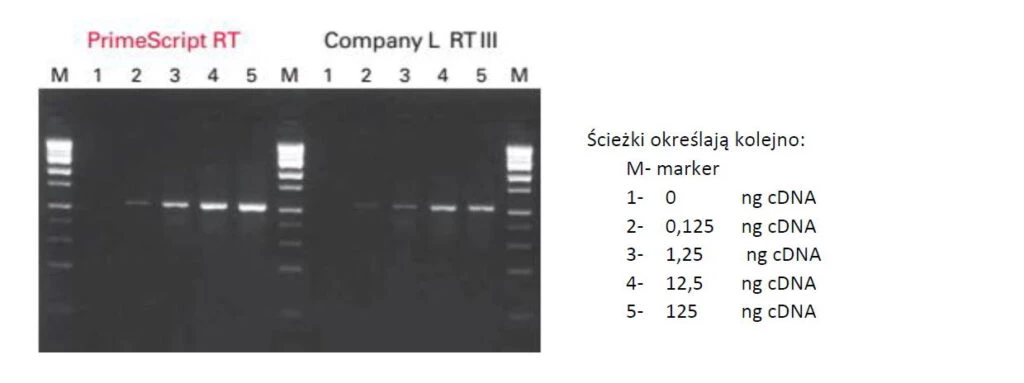
Fig.4 PrimeScript™ przepisze każdą matrycę.
Elektroforegram przedstawia rozdział produktów PCR dla fragmentu mRNA składającego się w ~75% z par GC (APOE). W doświadczeniu najpierw przepisano całe RNA pochodzące z ludzkiego serca w oparciu o primer oligo dT. Następnie odpowiednio rozcieńczono i użyto do reakcji PCR. PrimeScript™ przepisał wiernie i wydajnie mRNA genu bogatego w pary GC bez konieczności wstępnej denaturacji.
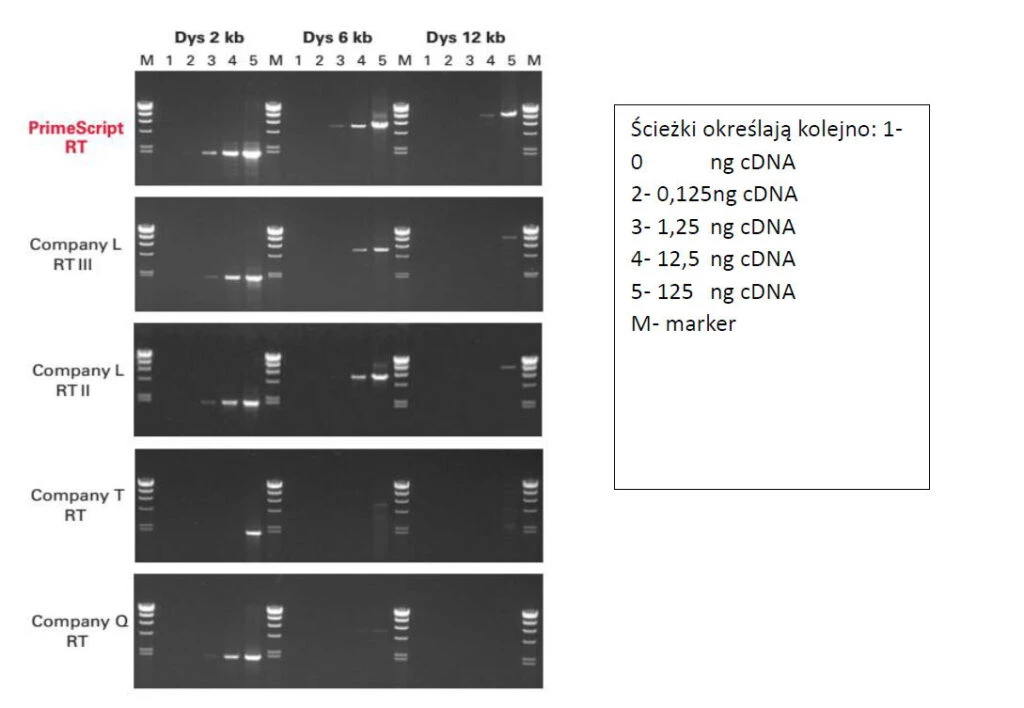
Fig.5 PrimeScript™ syntezuje długie cDNA. Elektroforegram przedstawia wyniki rozdziału trzech różnej długości odcinków przepisanego mRNA dystrofiny w różnych rozcieńceniach. cDNA do tej reakcji był przygotowany za pomocą czterech porównywanych odwrotnych transkryptaz i PrimeScript™. Wykorzystano primer oligo dT. Reakcję PCR przeprowadzono w takich samych warunkach dla wszystkich cDNA (po 30 cykli). PrimeScript™ przepisał najwydajniej RNA oraz wygenerował odcinki o długości 12 kb najefektywniej pośród porównywanych enzymów. Zdolność do tworzenia długiego cDNA jest istotna dla późniejszego etapu reakcji PCR. Tylko w pełni przepisany transkrypt jest gwarancją syntezy specyficznego amplikonu.
Ponad to przepisuje także długie matryce - 12 kb (Fig.5)! Mamy pewność, że z każdego przyłączonego primery oligo dT czy heksameru zostanie przepisana cząsteczka pełnej długości.
Jeśli już wspominamy o heksametrach – warto się zastanowić jak reakcja odwrotnej transkrypcji wygląda kiedy znajdują się one na nici RNA tworzą struktury dwuniciowe tzw. double stranded (ds).
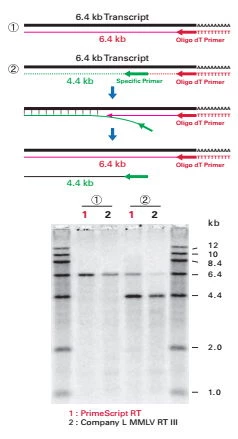
Fig 6. Zdolności do rozplatania nici przez PrimeScript™.
Reakcje przeprowadzono z udziałem PrimeScript™ i innego enzymu MMLV RT III. Zaprojektowano dwa warianty doświadczalne przedstawione schematycznie na rysunku. W oznaczonym jako pierwszy przeprowadzano klasyczną transkrypcję matrycy w oparciu o primer oligo dT. W jej wyniku powstawał odcinek 6,4 kb (a.). W drugim wariancie posłużono się tą samą matrycą i primerem oligo dT, ale mieszaninę wzbogacono o primer genospecyficzny. Przepisany na jego podstawie odcinek powinien mieć długość 4,4 kb. Podczas tej reakcji, kiedy następowała synteza z primera genospecyficznego, matryca była niejako blokowana. Transkrypcję w oparciu o primer oligo dT wstrzymywał przyłączony primer genospecyficzny. W ten sposób stworzono sytuację zbliżoną do tej, w której RNA formuje strukturę wyższego rzędu. Enzym bez zdolności do rozplatania nici nie jest w stanie przepisać matrycy od końca czapeczki poli A, a więc wygenerować dwóch fragmentów. PrimeScript™, dzięki swoim specjalnym zdolnościom do rozplatania nici, przepisał RNA z obu primerów.
Klasyczne odwrotne transkryptazy po napotkaniu przeszkody na swojej drodze odpadają od nici – jak przedstawia to eksperyment z Fig.6. Widzimy, że konkurencyjny enzym bez właściwości strand displacement, nie przepisywał dłuższego fragmentu mimo iż w 1 wariancie eksperymentalnym generował produkt bez problemu. PrimeScript przepisywał matryce nie zależnie od użytego primera (genospecyficznego produkt 4,4 kb czy oligo dT 6,4 kb). Było to możliwe ponieważ wnikał pomiędzy nici na odcinku gdzie wcześniej doszło do syntezy z primera genospecyficznego. Kiedy pracujemy z losowymi heksamerami co chwila tworzą się struktury ds. Enzymy odpadają od nici i nasze biblioteki rożnią się z przepisania na przepisanie. Dzięki właściwością PrimeScript każdy przyłączony heksamer skończy się przepisaniem pełnego transkryptu. Otrzymujemy biblioteki najbardziej odpowiadające faktycznej biologicznej reprezentacji transkryptów i bardziej powtarzalne z eksperymentu na eksperyment. To istotne zwłaszcza w przypadku porównywania wariantów eksperymentalnych w analizie ekspresji – RT-qPCR.
Wygenerowane cDNA może jednak służyć także jako insert przy klonowaniu. Ważna jest wtedy nie tylko długość i kompletność wygenerowanych fragmentów ale także wierność biologiczna. PrimeScript™ jest stosunkowo wiernym enzymem (Fig.7).
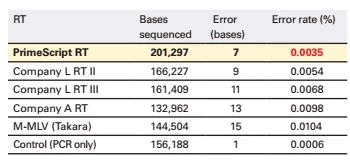
Fig.7 PrimeScript generuje wierne matryce. Tab. przedstawia wyniki sekwencjonowania cDNA wygenerowanego przez 5 różnych odwrotnych transkryptaz. PrimeScript™ popełnił jedynie 7 błędów na 201 tyś pz. uzyskując najlepszy wynik wierności spośród porównywanych enzymów. Poprawność odwrotnej transkrypcji jest kluczowa dla późniejszej, wydajnej reakcji PCR i bezbłędnego oddziaływania matrycy cDNA z primerami.
Wierność matrycy jest także istotna przy rozpatrywaniu efektywności reakcji qPCR, kiedy zależy nam na faktycznym pomiarze ilości mRNA (https://biokom.com.pl/?baza-wiedzy=real-time-pcr ).
Strand displacement to także szybkość pracy – cDNA do RT-qPCR w 15 min!
Z racji na różne wymagania odnośnie generowanego cDNA, zestawy do RT-qPCR różnią się od tych do RT czy RT-PCR w układzie end point. W tych drugich temp. reakcji jest podwyższona do 42oC aby umożliwić np. przyłączanie primerów genospecyficznych. PrimeScript™ dostępny jest w różnych zoptymalizowanych formatach.
| Aplikacja | qPCR | |||||
| Two-Step RT-qPCR | One-Step RT-qPCR (primery genospecyficzne do RT) |
|||||
| Nazwa | PrimeScript™ RT Master Mix (Perfect Real Time) |
(losowe heksamery i primery oligo dT) | PrimeScript™ RT Reagent Kit with gDNA Eraser (Perfect Real Time) |
One Step PrimeScript™ RT-PCR Kit (Perfect Real Time) |
One Step PrimeScript™ III RT-qPCR Mix (Z/bez UNG) |
One Step TB Green™ PrimeScript™ RT-PCR Kit II (Perfect Real Time) |
| Opis | Kompletny MasterMix | Elastyczny Reagent Kit |
Kit umożliwiający usunięcie gDNA | One-Step na sondy | One-Step na sondy, odporny na zanieczyszczenia | One-Step na TB Green (SYBR Green) |
| # Kat. L. rxns | RR036A 200 rxn RR036B 800 rxn (10 µL) |
RR037A 200 rxn RR037B 800 rxn (10 µL) |
RR047A 100 rxn RR047B 400 rxn (20 µL) |
RR064A 100 rxn (50 µL) |
RR601A 200 rxns (UNG) RR601B 1000 rxns (UNG) RR600A 200 rxns RR600B 1000 rxns (25 ul) |
RR086A 100 rxn (50 µL) |
| Skład | PrimeScript™ RT Master Mix (PrimeScript™, RNase Inhibitor, Oligo dT Primer, Random Hexamers, dNTPs, reaction buffer plus) RNase Free H2O EASY Dilution |
PrimeScript™ Buffer with dNTPs PrimeScript™ RT Enzyme Mix I with Rnase inhibitor Oligo dT Primer Random Hexamers RNase Free H2O EASY Dilution |
gDNA Eraser gDNA Eraser Buffer PrimeScript™ RT Enzyme Mix I with Rnase inhibitor PrimeScript™ Buffer 2 with dNTPs RT Primer Mix (Oligo dT Primer & Random Hexamers) RNase Free H2O EASY Dilution |
One Step RT-PCR Buffer III with dNTPs TaKaRa Ex Taq HS PrimeScript™ RT enzyme Mix II with Rnase inhibitor RNase Free H2O ROX I & II Reference Dye |
One Step PrimeScript III RT-qPCR Mix, with/without UNG (2X) RNase Free H2O ROX Reference Dye ROX Reference Dye II |
One Step SYBR RT-PCR Buffer with dNTPs & TB Green PrimeScript™ 1 step Enzyme Mix 2 with TaKaRa Ex Taq HS & Rnase inhibitor RNase Free H2O ROX I & II Reference Dye |
| Aplikacja | End-Point PCR Synteza cDNA |
End-Point PCR Synteza cDNA |
End-Point PCR Synteza cDNA |
End-Point PCR Two-Step RT-PCR |
End-Point PCR Two-Step RT-PCR |
End-Point PCR One-Step RT-PCR (primery genospecyficzne do RT) |
End-Point PCR One-Step RT-PCR (primery genospecyficzne do RT) |
| PrimeScript™ Reverse Transcriptase | PrimeScript ™ 1st strand cDNA synthesis kit | PrimeScript IV 1st Strand cDNA synthesis mix | PrimeScript™ RT-PCR Kit | PrimeScript™ High Fidelity RT-PCR Kit | PrimeScript™ One Step RT-PCR Kit Ver. 2 | PrimeScript™ One Step RT-PCR Kit, Ver.2 (Dye Plus) | |
| Opis | Podstawowa odwrotna transkrypcja | Tworzenie biblioteki cDNA | Tworzenie biblioteki cDNA; szybko i bez grzania próbki |
Tworzenie biblioteki cDNA I amplifikacja targetu | Tworzenie biblioteki cDNA I wierna amplifikacja targetu | One-Step w układzie end point | Wygodny One-Step w układzie end point z dodanym barwnikiem i obciążnikiem |
| # Kat. L. rxns | 2680A 10 000 U (100 rxns; 20 µL) | 6110A 50 rxn (20 µL) | 6215A 50 rxns (20 ul) |
RR014A 50 rxn (20 µL RT; 50 µL PCR) | R022A 50 rxn (20 µL RT; 50 µL PCR) | RR055A 50 rxn (50 µL) | RR057A 50 rxn (50 µL) |
| Skład | PrimeScript™ Rtase PrimeScript™ Buffer | PrimeScript™ Rtase PrimeScript™ Buffer RNase Inhibitor dNTP Mixture Oligo dT Primer Random Hexamers RNase Free H2O | 5X PrimeScript IV cDNA Synthesis Mix zawierający primer oligo dT, PrimeScript IV RTase, RNase Inhibitor, dNTP Mixture I Reaction Buffer (with Mg2+) Random 6 mers RNase Free H2O | PrimeScript™ Rtase PrimeScript™ Buffer RNase Inhibitor dNTP Mixture Oligo dT Primer Random Hexamers TaKaRa Ex Taq HS PCR Buffer II . Positive RNA Control F-1 & R-1 Control Primer RNase Free H2O | PrimeScript™ Rtase PrimeScript™ Buffer RNase Inhibito dNTP Mixture Oligo dT Primer Random Hexamers PrimeSTAR Max Premix . Positive RNA Control Control Primer F-1 & R-1 RNase Free H2O | PrimeScript™ 1 step Enzyme Mix with TaKaRa Ex Taq HS and Rnase inhibitor 1 step Buffer (Enhancer & dNTPs Plus) Control Primer F-1 & R-1 Positive RNA Control RNase Free H2O | PrimeScript™ 1 step Enzyme Mix with TaKaRa Ex Taq HS and Rnase inhibitor 1 step Buffer (Dye, Enhancer & dNTPs Plus) Control Primer F-1 & R-1 Positive RNA Control RNase Free H2O |





